Rev 01/27/26
Dorylaimida Pearse, 1942
There are two cuticle types in the Dorylaimida: (a) dorylaimoid with the inner layer not loose or separated from the outer layes and without radial cuticular elements; (b) tylencholaimoid in which the inner cuticular layer is loose and there are irregular radial elements. Among all the families of the Dorylaimida, only Leptonchidae and Tylencholaimidae have the tylencholaimoid cuticle; other families have a dorylaimoid cuticle (Wu et al., 2019).
The first species in the Dorylaimida group (Dorylaimus stagnalis) was described by Dujardin in 1845. The order Dorylaimida was proposed by Pearse in 1942.
The Order Dorylaimida is probably the most speciose and diverse of all nematode taxa; the soil forms are among those most poorly studied.
Key to the Order Dorylaimida (suborders, superfamilies, families and genera)
Classification and Characteristics Charts of the Orders Dorylaimida and Triplonchida
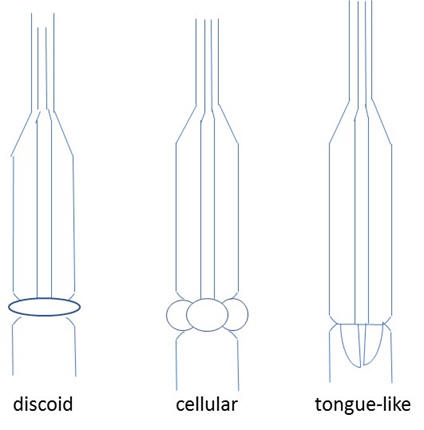
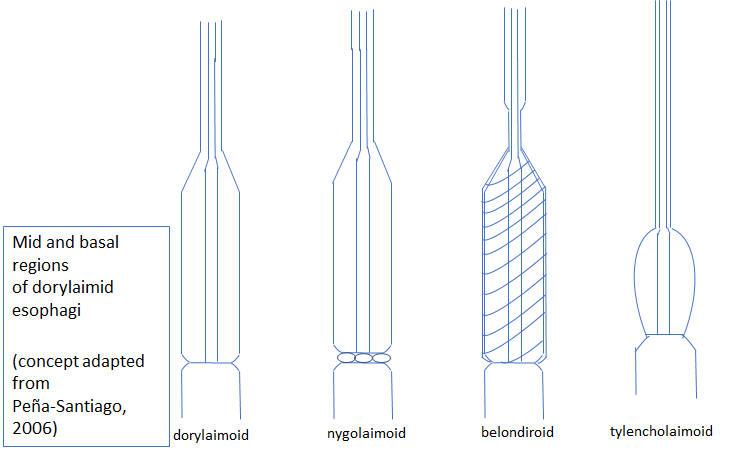
Some dorylaimid esophago-intestinal cardia configurations
Although the tail region of dorylaims varies in length and shape, three basic forms can be defined:
1. filiform tails that are usually > 3x the anal/cloacal body diameter. (a/cbd)
2. conical tails that are short, usually < 3x a/cbd
3. rounded tails that are very short, usually < 1x a/cbd.
There are also many varioants to these general categories: cylindrical, clavate, digitate, etc.
Among the three major tail shape categories, three postembryonic developmental patterns can be identified:
a) tail shape is generally similar in males and females and does not exhibit sexual; dimorphism. In genera of the Dorylaimidae without sexual dimorphism of tail shape (adults of both sexes with long tails), there is no change in tail shape during development. Similarly, in the conical-tailed Qudsianematidae, tail shape does not change during developmental stages.
b) in some genera females have filiform tails and males have rounded tails. Such sexual dimorphism occurs in some genera in the family Dorylaimidae whereas it does not occur in the family Qudsianematidae where tail shape in both males and females is the same and either conical of rounded.
c) in some rounded-tail Qudsianematidae, juvenile stages have pointed tails that become shorter and rounded at specific molting events.
Plant-parasitic nematodes in the Dorylaimida are ectoparasites. They
exhibit some adaptations to greater productivity in their energetics and
influence on the host:
1. Immobilize
feeding site (root tip)
2. Deeper tissue
penetration
3. Nurse cell
modifications - increased cell surface increased metabolic activity
Additionally, they are the group of nematodes
that transmit plant viruses: NEPO
(nematode-transmitted polyhedral) viruses by the Longidoridae. Examples: tomato
ringspot, grapevine fanleaf
Note: in 1995 DeCraemer moved the Trichodoridae to the Triplonchida on the basis of odontostyle and esophagus morphology and ultrastructure of the cuticle.
Not all Dorylaimida have a stylet (sometimes called spear to distinguish it from the stomatostyle of the Tylenchida). Some have a tooth. In the few Dorylaimida specialized for plant parasitism, the spear is an odontostyle. In some predatory Dorylaimida the tooth, positioned subventrally in the stoma, lacks a lumen. When prey are punctured, food passes along the tooth into the stoma. In other Dorylaimida, including plant parasites and predators such as Labronema, the spear is hollow and food passes through its lumen. The lumen is formed by longitudinal infolding which is marked by a seam along its dorsal surface. Where there is a spear extension or odontophore, it is a specialization of the cuticle-lined esophageal lumen. The odontostyle and odontophore are protracted and retracted as one unit respectively by sets of eight and four muscles (Baldwin et al, 2004).
Baldwin, J.G., S.A. Nadler and B.J. Adams. 2004. Evolution of plant parasitism among nematodes. Ann. Rev. Phytopathol, 42:83-105.
Dujardin, F. 1845. Histoire Naturelle des Helminthes ou Vers Intestinaux. Paris. 654p.
Jairajpuri, M.S. and W. Ahmad. 1992. Dorylaimida: free-living, predaceous and plant-parasitic nematodes. Brill, Leiden. 458 p.
Pearse, A.S. 1942. Introduction to Parasitology. Baltimore. 357p.
Pena-Santiago, R. 2006. Dorylaimida Part 1. Superfamilies Belondiroidea, Nygolaimoidea and Tylencholaimoidea. Pp 326-391 in E. Abebe, István Andrássy, W. Traunspurger (eds) Freshwater Nematodes: Ecology and Taxonomy. CABI Publishing.
Pena-Santiago, R. and S. Alvarez-Ortega. 2014. An integrative approach to assess the phylogeny and the systematics of rounded-tailed genera of the subfamily Qudsianematinae (Nematoda, Dorylaimida). Zoologica Scripta 43:418-428.
Wu, W-J., Xu, C-L., Xie, H., Wang, D-W. 2019. Three new species, one new genus and subfamily of Dorylaimida (de man, 1876) Pearse, 1942, and revisions of the families Tylencholaimellidae Jairajpuri, 1964 and Mydonomidae Thorne, 1964 (Nematoda, Dorylaimida). PeerJ 7:e7541 https://doi.org/10.7717/peerj.7541