Family HETERODERIDAE
Classification:
Chomadorea
Chromadoria
Rhabditida
Tylenchina
Tylenchoidea
Heteroderidae
Filip'ev & Schuurmans Stekhoven, 1941
Synonyms:
Meloidogynidae (Skarbilovich, 1959)
Meloidoderidae (Golden, 1971)
Ataloderidae (Wouts, 1973)
Nematode genera in the family Heteroderidae can be divided into two
major groups: the cyst nematodes in which the famale body becomes a hard-walled
cyst, and the cystoid nematodes in which the body wall of the female does not
harden (Subbotin et al., 2017).
- Sedentary obligate parasites of roots, forming galls in some cases.
- Marked sexual dimorphism, body vermiform and slender in second-stage juveniles, robust in males, and inflated in females. Labial region and cephalic framework well developed, especially in males and juveniles, secondarily reduced in females.
- Stylet robust; cone half the length of total stylet.
- Dorsal esophageal gland opening close to base of stylet.
- Median esophageal bulb usually large, with strong valves.
- Esophageal glands overlapping intestine ventrally and also laterally.
Female:
- Sedentary in root tissues.
- Globose (exception: Verutus with sausage-shaped females).
- Vulva generally terminal or subterminal (exception:
Verutus and Meloidodera where equatorial). Two genital branches,
amphidelphic or prodelphic. Columned uterus with three rows of cells.
- Eggs laid in a gelatinous matrix (exception: Verutus) or totally or partially retained within female body of which the cuticle may be transformed (cysts).
Male:
Second stage juveniles:
- Tail conical, with long hyaline posterior part.
- Phasmids anterior to half tail length.
Two sub-families:
| Characteristic |
Heteroderinae |
Meloidogyninae |
| Feeding site |
Multinucleate syncytium |
Multinucleate giant cell |
| Host range |
Narrow |
Wide |
| Reproductive strategies |
Sexual |
Mainly parthenogenic |
| Eggs |
Mainly retained in female body |
Deposited in egg mass |
| Female body |
Becomes hardened cyst |
Does not form cyst |
| Hatching factors |
From host root exudates |
Favorable environmental conditions. |
| Root penetration |
Inter- and intracellular migration directly across cortex to
zone of cell differentiation. |
Longitudinally, mainly intercellularly, toward root tip and
then turning into zone of differentiating cells. |
General Characteristics of cyst nematode genera
Large sectors of the developing root, including areas that would have become
vascular tissue are transfomed into syncytia (Heteroderidae) or giant cells
(Meloidogynidae). Syncytia and giant cells have many plastids,
mitochondria, ribosomes, increased rough endoplasmic reticulum and enlarged
lobed nuclei.
Cell wall protruberances increase the surface area of the cell membrane for
flow of solutes from the xylem to the syncytium - the transfer cell
configuration (Endo, 1975).
More than 50 genes are upregulated to some extent in the
development of giant cells (Meloidogyne) and syncytia (Heterodera/Globodera) (Gheysen and Fenoll, 2002). Both types
of feeding cells have the genome amplified as a result of multiple shortened
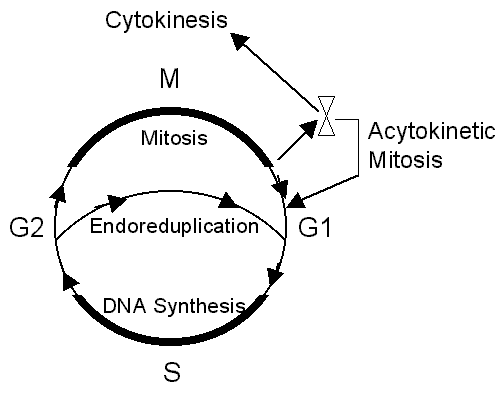
cell cycles; but the processes differ. Giant-cells go through repeated (acytokinetic)
mitosis. Syncytia undergo repeated S-phase endoreduplication
without mitosis or nuclear division.
Since nematodes in the Heteroderidae become sedentary from the late second
stage onwards (except for the metamorphosis to males), the feeding site in the
plant must be maintained in a condition favorable for perhaps five or six weeks
to allow the nematode to fulfill its reproductive potential. Besides
stimulation of the cell cycle events,
pathogen-triggered immunity
(PTI) responses, including activation of the salicylic acid pathway, must be
suppressed. The salicylic acid pathway leads to production of active oxygen
molecules and hypersensitive cell death. In the Meloidogynidae, a possible
candidate for effector-triggered suppression of PTI is chorismate mutase,
produced in the nematode esophageal glands. In PTI responses, chorismate
is converted to salicylic acid to iniate the defense events.
Chorismate mutase from the nematode reduces chorismate, and thus
salicylic acid (Smant and Jones, 2011). In Heteroderinae, the Hg30C02 effector protein of
Heterodera
glycines may be involved in active
suppression of host defenses. The same gene occurs in H. schachtii but
not in Meloidogyne spp. (Hamamouch et al., 2012).
As of 2013, some 70 proteins, many new or unknown, have been detected in
nematode secretions into plant cells (Thomas Baum, pers. com.)
References:
Return to Heteroderidae
Menu
Go to Nemaplex Main Menu