|
 |
Rev 10/30/2019
|
|
|
Terrestrial plants are
estimated to have annual production of 120 billion tons of
biomass - 5% minerals = 6 billion tons of minerals mined from the soil each year.
A plant may transpire its own weight in water in a day.
The integrity of Casparian strip (waxy layer around endodermis cells) is important - nutrients, and sometimes water, are taken up against a gradient.
Secondary roots and endoparasites are disruptive to the Casparian strip.
Total energy consumption during the lifecycle of a female root-knot nematode (Meloidogyne spp.) is 1 calorie.
The total biomass of a female root-knot nematode is 200 µg, including the egg mass (Melakerberhan and Ferris). For say 100,000 nematodes in a root system, the total nematode biomass is 20 g! Allowing for 50% production efficiency, total material extracted from the plant would be 40 g. So, the demand effect on the plant may be minimal unless plant is very stressed and resources are limited.
An adult Heterodera schachtii consumes 11 nL/day of cell content (Muller et al, 1981). So, it would take 1,000,000 such females to remove 11 ml of cell content in a day.
a. Penetration of cells - relative to length of stylet. Damage will depend on types of cells affected - storage tissues, cortex, or functional vascular.
b. Migration through tissues - intercellular and intracellular requiring dissolution of cell walls, middle lamellae. Suggests cellulase and pectinase enzymes - spongy tissues, sloughing, e.g. damage caused by Pratylenchus and Ditylenchus. Allows ingress of other organisms. Root-knot (Meloidogyne spp.) and cyst (Heterodera spp.) produce endogluconase (cellulase) enzymes and pectate lyase which are presumable involved in the passage through plant tissues.
c. Leakage from damaged tissues - it is estimated that up to 20% or more of photosynthate partitioned to roots may leak into rhizosphere soil without root damage. "Root exudation" nurturing organisms in rhizosphere - presumably to plant benefit - but speculate that selection has optimized the costs and benefits. Enhancing root leakage through nematode damage must reduce plant productivity.
a. Nematode secretions - associated with establishment and maintenance of feeding sites. Effects increase with sedentary endoparasitism. Secretions from the nematode digestive glands may polymerize into a feeding tube inside the cell. The feeding tube remains associated with the stylet during ingestion. When the stylet is withdrawn the opening in the cell wall is sealed with an electron-dense feeding plug.
b. Physiological effects -
c. Whole-plant effects -
Disturbance of the biochemical
network. Wallace (1987) points to the complexity of the
biochemical pathways:
Photosynthesis divided into two basic phases - a light phase when
light energy is converted into chemical energy, and a synthetic
phase in which carbohydrates are formed in a series of reactions
accelerated by light. Photosynthesis involves a chain of
metabolic events cross-linked to other physiological processes,
so disruption of one may have effects throughout system.
For example, Bird suggested that photosynthesis is reduced in
tomato by
Meloidogyne
javanica by
inhibiting production of cytokinins and gibberellins in roots,
and/or by increased stomatal resistance due to water stress.
Fatemy et al. indicate that the response of potato to
Globodera
rostochiensis is due to stomatal closure through water
stress; the result is reduced photosynthesis.
However, generally the mechanisms by which root-infecting
pathogens, including nematodes, affect physiological processes
have been insufficiently studied.
d. Plant as an Integrator - Metabolic pool concept - plant as an integrator - concepts of demand and damage. Melakeberhan and Ferris characterized five effects of root-knot nematode infection in grape while exploring the impact in an energy partitioning and flow model:
Successful Parasitism by Plant-parasitic Nematodes
Suppression of Plant Defenses by Nematodes
Biotrophic Pathogens
Pathogens that can only feed on a living host and must keep the host and its cells alive are termed biotrophic pathogens.
Sedentary ecto- and
endoparasitic nematodes are in this category, for example, species of
Meloidogyne, Heterodera, Xiphinema, Tylenchulus, Rotylenchulus.
Nematodes that withdraw contents form individual
cells and then move to new feeding sites are considered cell grazers.
Most plants are resistant
to most pathogens; they have highly effective immune systems.
Host defense mechanisms may be as extreme
as programmed cell death, the hypersensitive response.
All biotrophic pathogens must suppress host defenses. The feeding site must be induced without host detection or without induction of host defenses.
Following establishment of the feeding site by sedentary nematodes, it must be maintained for up to 5 or 6 weeks to allow the nematode to achieve its reproductive potential.
That time scale is much greater than that required by many bacterial and fungal pathogens of plants.
Failure to establish and
maintain the feeding site may prevent reproduction and therefore is
catastrophic to the nematode genotype.
Consequently, there is strong selection pressure on
nematodes to suppress host defenses.
Pre-existing Defenses – Basal Resistance
a.
Structural – cuticle,
wax, wall thickness, spines that suppress penetration of cells.
b.
Chemical-phenolic and
other compounds that inhibit or kill invading organisms.
PAMP Signals
Organisms attempting to feed on plant cells, or to invade plant tissues, betray their presence with recognizable molecular signals on their surfaces.
Such signals are termed pathogen associated molecular patterns (PAMPs) and are recognized by pattern recognition receptors on cell surfaces.
Chitins in fungal cell walls are PAMPS which trigger immunity responses (pathogen-triggered immunity – PTI).
The PAMP signals of nematodes are unknown; chitin is not present in the cuticle although it does occur in egg shells and perhaps in the stylet.
Plants characteristically deposit callose to strengthen cell walls at the point of invasion, including at the point of nematode stylet insertion.
Also, pathogen invasion
trigger the triggers jasmonic acid signaling pathway which stimulates
production and release of other defense toxins.
b. DAMP Signals
Another set of signals that may trigger PTI responses in
plants are cell-degradation products resulting from damage caused by the
invasion, damage-associated molecular patterns (DAMPs).
Invading bacteria and fungi, and probably nematodes, release effector molecules into plant cells to suppress PTI and render the plant susceptible to infection or invasion.
PAMP-triggered PTI, the first line of defense, may involve production of salicylic acid (SA) as a signal to invoke defense mechanisms. In that case, successful nematode infections would involve suppression of SA production, reduction of callose thickening of cell walls and suppression of active oxygen defense responses (H2O2, superoxide) which may initiate localized programmed cell death – hypersensitive response.
SA signaling is possibly
disrupted by chorismate mutase produced in the esophageal glands.
In the PTI signaling pathway, chorismate is converted to salicylic acid.
Chorismate mutase from the nematode reduces chorismate and thus SA, so
defense mechanisms are not triggered.
Incidentally, like cellulases, chorismate mutase is an example of
horizontal gene transfer from bacteria. Nematodes are the only metazoan with the
enzyme.
An alternative mechanism of PTI suppression by nematodes
is the production of effectors which cause ubiquitin to attach to plant signal
proteins and thus reduce their levels and effectiveness in triggering PTI
responses.
The evolution of effector suppression of PTI has resulted in evolution of immune receptors, with a nucleotide-binding domain and a leucine-rich domain (NB-LRR), in plants that recognize the effector molecules and activate effector-triggered immunity (ETI). However, successful pathogens have evolved next-generation effectors that suppress ETI.
One possible candidate is the Hg30C02 effector protein of Heterodera glycines which may be involved in active suppression of host defenses (Hamamouch et al., 2012).
Another is the 8D05 parasitism gene of Meloidogyne incognita which is required for successful infection of host roots. The gene codes for a protein that is secreted from the subventral glands duriing initiation of the feeding site (Xue et al., 2013).
Plants have responded with more specific ETIs and the evolutionary treadmill continues.
PTI responses to PAMPs and DAMPs are relatively general in their effect but higher level ETIs are progressively more specific to individual pathogens.
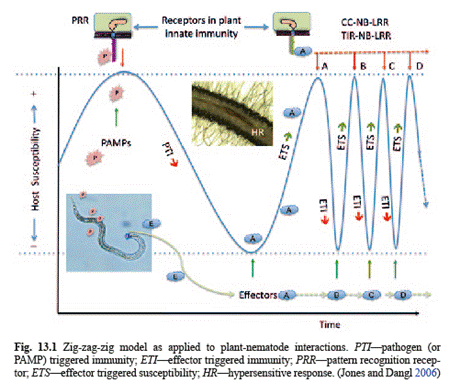
The cyclical evolutionary
process of plant-nematode interactions with regard to plant immunity and
susceptibility is depicted by the zig-zag-zig model (Jones and Dangl, 2006).
Initially PAMPs trigger PTI which reduces
susceptibility.
Then nematodes develop effectors that suppress
PTI and plants evolve immunity responses to the effectors.
|
|
The Evolutionary Response:
Effector-triggered Immunity (ETI)
In effect, the sources of specific ETIs are resistance genes. Thus, the Mi gene of tomato codes for receptors to the effector molecules introduced by root-knot nematodes to suppress plant defenses and, perhaps, facilitate the development of feeding sites.
However, although they are known for some other pathogens, the nematode effector molecules that trigger ETIs have not yet been determined and are the focus of several active research programs.
Although the nature of
nematode effector products has still to be determined, the hypersensitive
response of cells to the activated ETI effectively disrupts the feeding and
development of sedentary endoparasitic nematode species.
Suppression and Avoidance of Host Defenses
Glutathione peroxidases on surface coats reduce active
oxygen plant defenses.
Many plant-parasitic nematodes produce glutathione S
tranferases that detoxify endogenous toxic molecules.
They also produce superoxide dismutase that breaks down active oxygen
plant defenses.
|
|
 |
|
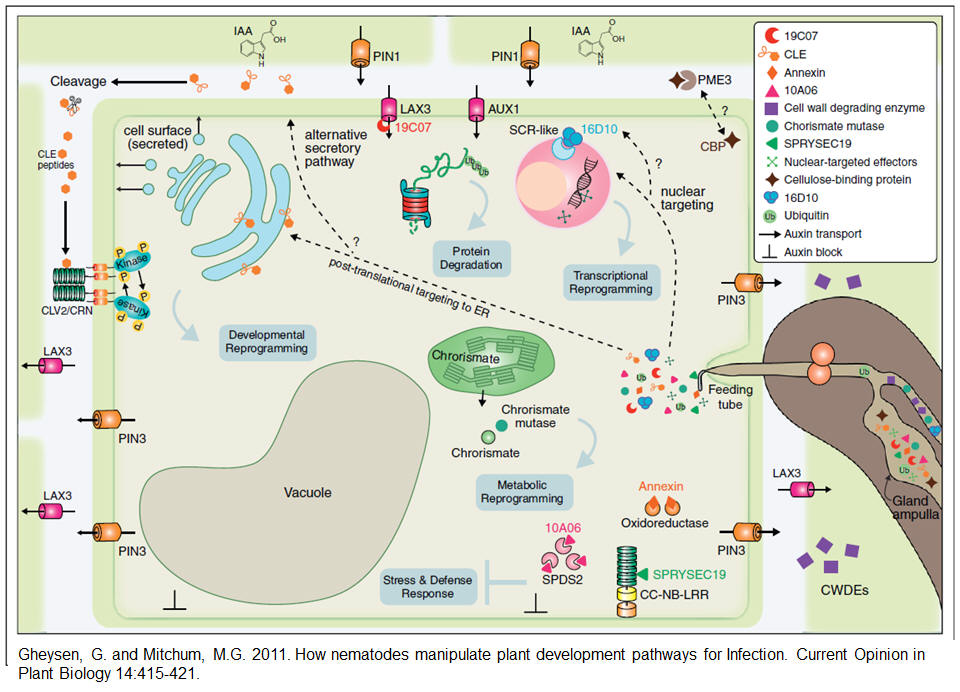
The above compilation by Gheysen and Mitchum (2011) is based mainly on research on cyst and root-knot nematodes.
Cell wall degrading enzymes (CWDEs) and chorismate mutase are secreted by both root-knot and cyst nematodes while 16D10 is specific for root-knot nematodes. Effector proteins originating from the subventral and dorsal esophageal gland cells are secreted into plant tissues through tthe nematode stylet.
Synopsis from Gheysen and Mitchum (2011). |
The genome of plant-feeding nematodes of the sub-order Tylenchina includes genes that encode for endoglucanases. Endogluconases are cellulases, a family of enzymes formerly thought to be restricted to prokaryotes. Other plant-cell wall digesters such as termites and ruminants use symbiotic and commensal bacteria to achieve dissolve cellulose. The presence of these and other genes suggests that horizontal or lateral gene transfer has occurred between bacteria and nematodes.
Koch, Pasteur - the germ theory - required rules of proof.
Mountain provided guidelines for obligate parasite nematodes:
Note - term "interaction" is loosely used - implies that effect in combination is different than sum of individual effects - not additivity. Three descriptions of the result of combinations of organisms: -synergistic, -suppressive, -no interaction.
- Reduced tolerance - multiple stress
- Vectoring - Longidoridae and Trichodoridae
- Change in substrate - fungi?
- Routes of ingress - fungi, bacteria
- Leakage - energy source - fungi, bacteria
- Induced resistance - any examples?
- Reduction of stress - mycorrhizae
- Biological antagonists - but not an interaction of two pathogens
- Reduced substrate availability
See Sasser and Freckman
in Vistas on Nematology.
Questionnaires returned by 371 nematologists worldwide (handout
with Tables 2 and 3)
Consider inherent biases in data of this kind.
Consider means and variance in crop loss data.
- almost nobody had 10% yield loss
- include management costs as part of the loss
Ranking of important genera and relative weight:
|
Note - considerable variation by region, so questionnaire data biased by number of respondents per region.
International Survey of Crop Losses due to Nematodes
|
Life-sustaining Crops |
Annual Loss (%) |
Economically-important Crops |
Annual Loss (%) |
|
19.7 |
10.5 |
||
|
6.3 |
4.2 |
||
|
8.4 |
15.0 |
||
|
13.7 |
10.7 |
||
|
17.1 |
15.1 |
||
|
10.2 |
Eggplant |
16.9 |
|
|
10.9 |
Forages |
8.2 |
|
|
11.8 |
12.5 |
||
|
4.2 |
Guava |
10.8 |
|
|
12.0 |
Melons |
13.8 |
|
|
13.2 |
Misc. other |
17.3 |
|
|
12.2 |
Okra |
20.4 |
|
|
10.0 |
Ornamentals |
11.1 |
|
|
3.3 |
Papaya |
15.1 |
|
|
6.9 |
12.2 |
||
|
10.6 |
14.9 |
||
|
10.9 |
8.2 |
||
|
15.3 |
14.7 |
||
|
Sweet potato |
10.2 |
20.6 |
|
|
7.0 |
Yam |
17.7 |
|
|
|
|
|
|
|
Average |
10.7% |
Average |
14.0% |
|
|
Overall Average 12.3% |
|
|
|
|
|
|
|
|
Information based on a worldwide survey with 371 responses. Source: Sasser, J.N., Freckman, D.W., 1987. A world
perspective on Nematology: the role of the society.
Pp 7-14 in J.A. Veech and D.W. Dickson (eds) Vistas on Nematology.
Society of Nematologists, |
|||
Estimated Crop Losses 2008
| Crop | Number of estimates per crop | FAO production estimates (1000 metric tons) | Estimated price per metric ton -2008 (U.S.$) | Estimated yield losses due to nematodes (%) | Estimated monetary loss due to nematodes - 2008 (x1000 U.S.$) |
| Banana | 78 | 81,263 | 928 | 19.7 | 14,855,056 |
| Barley | 49 | 136,209 | 238 | 6.3 | 2,044,978 |
| Cassava | 25 | 228,138 | 175 | 8.4 | 3,353,629 |
| Citrus | 102 | 105,000 | 711 | 14.2 | 10,601,170 |
| Cocoa | 13 | 4,012 | 2,693 | 10.5 | 1,134,626 |
| Coffee | 36 | 7,742 | 1,915 | 15 | 2,223,425 |
| Corn | 125 | 637,444 | 183 | 10.2 | 11,895,929 |
| Cotton (lint only) | 85 | 112 | 1,040 | 10.7 | 12,463 |
| Field bean | 70 | 6,371 | 1,200 | 10.9 | 833,327 |
| Oat | 37 | 25,991 | 117 | 4.2 | 127,327 |
| Peanut | 69 | 30,670 | 1,470 | 12 | 5,410,188 |
| Potato | 141 | 321,736 | 264 | 12.2 | 10,362,473 |
| Rice | 64 | 432 | 624 | 10 | 26,957 |
| Sorghum | 53 | 64,589 | 59 | 6.9 | 262,942 |
| Soybean | 91 | 56,389 | 339 | 10.6 | 2,024,967 |
| Sugar beet | 51 | 247,878 | 47 | 10.9 | 1,258,234 |
| Sugar cane | 65 | 1,557,664 | 36 | 15.3 | 8,462,835 |
| Sweet potato | 67 | 126,299 | 407 | 10.2 | 5,242,210 |
| Tea | 16 | 3,871 | 282 | 8.2 | 89,637 |
| Tobacco | 92 | 6,326 | 6,600 | 14.7 | 6,137,485 |
| Wheat | 89 | 676,300 | 237 | 7 | 11,237,807 |
| Sources: | |||||
| http://www.fas.usda.gov/wap/circular/2008/08-09/productionfull09-08.pdf | |||||
| http://usda.mannlib.cornell.edu/usda/current/PeanPrice/PeanPrice-10-10-2008.pdf | |||||
| http://usda.mannlib.cornell.edu/usda/current/CropProdSu/CropProdSu-01-11-2008.pdf | |||||
| http://www.nass.usda.gov/Publications/Ag_Statistics/2008/index.asp | |||||
References:
Bird, A.F.
Davis. E.L., R.S. Hussey, T.J. Baum, J. Bakker, A. Schotts, M.N. Rosso and P. Abad. 2000. Nematode parasitism genes. Ann. Rev. Phytopathol. 38:365-396.
Dropkin, V.
Garrett
Hamamouch, N., Li, C., Hewezi, T., Baum, T.J., Mitchum,
M.G., Hussey, R.S., Vodkin, L.O., Davis, E.L. 2012.
The interaction of the novel Hg30C02 cyst nematode effector protein with a
plant b-1,3-endoglucanase may suppress host defence to promote parasitism.
Journal of Experimental Botany.
Smant, G., Jones, J. 2011.
Suppression of plant defences by nematodes.
Chapter 13, pp 273-286. In Jones, J., Gheysen, G., Fenoll, C. (eds).
Genomics and Molecular Genetics of Plant-Nematode Interactions.
Springer, NY.
Jones, J.D.G, Dangl, J.L. 2006. The plant
immune system. Nature 444:323-329.
Jones, J. 2012.
Lectures in the EUMAINE program, University of Ghent.
Gheysen, G. 1998. Chemical signals in the plant-nematode interaction. A complex system? In Romeo et al. Phytochemical signals and plant-microbe interactions.
Gheysen, G. and Mitchum, M.G. 2011. How nematodes manipulate plant development pathways for Infection. Current Opinion in Plant Biology 14:415-421.
Hussey, R.S. and V.M. Williamson 1998. Physiological and molecular aspects of nematode parasitism. Agronomy Monograph 36.
Koenning, S.R., Overstreet, C., Noling,, J.W., Donald, P.A., Becker, J.O., Fortnum B.A. 1999. Survey of Crop Losses in Response to Phytoparasitic Nematodes in the United States for 1994. Journal of Nematology 31:587-618.
Melakeberhan and Ferris
McClure
Muller et al, 1981
Sasser, J.N.,
Freckman, D.W., 1987. A world perspective on Nematology: the role of the
society.
Pp 7-14 in J.A. Veech
and D.W. Dickson (eds) Vistas on Nematology. Society of
Nematologists,
Seinhorst
Wallace, 1986
Wyss U., F.M.W. Grundler and A. Munch. 1992. The parasitic behaviour of second-stage juveniles of Meloidogyne incognita in roots of Arabadopsis thaliana. Nematologica 38:98-111.