Rev 01/27/26
Usually slender, elongate, small species. Sexes similar. Stylet usually small, delicate.
![]() photomicrograph
by Howard Ferris and Sam Woo, UC Davis.
photomicrograph
by Howard Ferris and Sam Woo, UC Davis.
Esophagus with slender procorpus, median bulb either well-developed, weakly developed or even not obvious; long, slender isthmus, esophageal glands symmetrically arranged, pyriform, rarely with slight overlap of intestine.
Female: One gonad, prodelphic; rarely, two gonads; columned uterus with four rows. Tail long, conoid to filiform.
Male: Caudal
alae adanal, small, occasionally lacking. 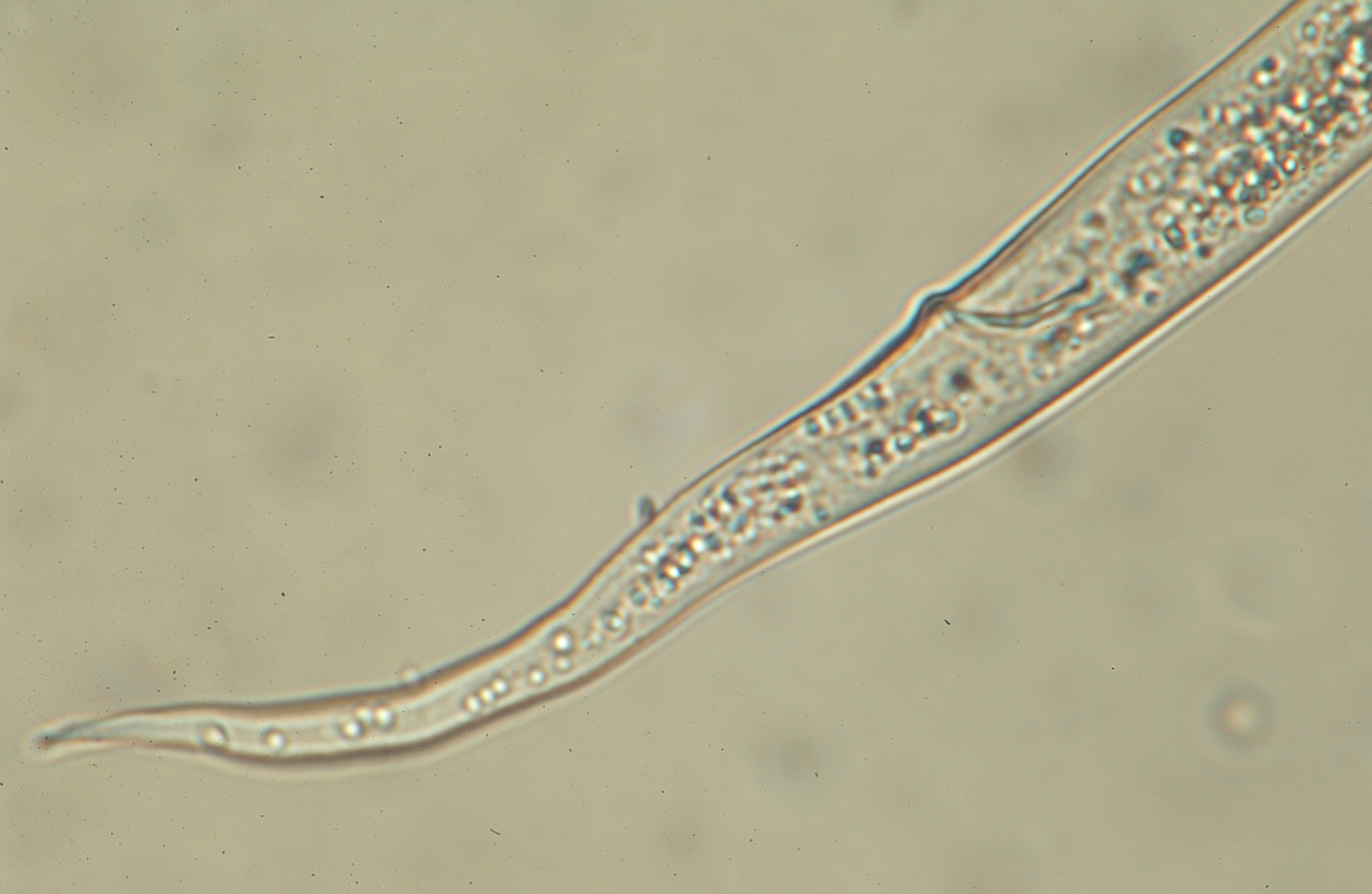
photomicrograph by Howard Ferris and Sam Woo, UC Davis
Most of the characteristics of the species of Tylenchidae conform with the definition of primitivity and as expressed by Luc et al. (1987): small stylet, delicate labial framework; amphid openings from small to elongate, sinuousoid or straight, extending posteriad longitudinally; median bulb spindle shaped, small, delicate or rounded, muscular; esophageal glands symmetrically arranged, pyriform; tail elongate, long many of which are generally considered to be ancestral characters (Qing and Bert, 2017)..
The family Tylenchidae is one of the most diverse and cosmopolitan of the Tylenchina; apparently actively evolving in some characters, such as the amphids moving from large elongate post labial structures, varying to arc-shaped or oval pits limited to the labial plate, or even small elliptical apertures near the oral opening (Miculenchus, Ecphyadophora), with many links and relationships with other groups (i.e. Tylodorus with characteristics of the Dolichodoridae; Filenchus with characteristics of the Anguinidae, etc.).
As of 2019, the family Tylenchidae contains over 400 species (Siddiqi, 2000; Geraert, 2008) with high ecological diversity and abundance. Many species are poorly described on the basis of morphological characters and a limited number of specimens. Molecular data are improving definition of species characteristics (Panahandeh et al., 2018).
The family is most closely related to the Anguinidae, which were formerly considered part of the Tylenchidae (Siddiqi, 1971). The families differ in that there are small elliptical amphids in Anguinidae, which also have elongated, axial spermatheca, large sperms with prominent cytoplasm, and females with a long post-uterine branch of the gonad (more than two body diameters at vulva level).
adapted from Geraert and Raski, 1987
| 1 | a | Females diovarial | 2 | |
| b | Females monovarial | 5 | ||
| 2 | a | Tail short, subcylindrical, rounded; stylet very long (90-110µm) | Macrotrophurus | |
| b | Tail elongate, attenuated; stylet < 20µm | 3 | ||
| 3 | a | Cephalic framework sclerotized; vulva with lateral membranes; male cloaca with hypoptygma | Antarctenchus | |
| b | Cephalic framework not sclerotized; vulva withithout lateral membranes; male cloaca without hypoptygma | 4 | ||
| 4 | a | Head high, amphidial slit obvious; metacorpus posterior to middle of esophagus | Psilenchus | |
| b | Head low, amphidial slit indistinct; metacorpus anterior to middle of esophagus | Atetylenchus | ||
| 5 | a | Stylet very long (76-104µm) | Tylodorus | |
| b | Stylet long (38-52µm) | Epicharinema | ||
| c | Stylet moderate (22-34µm) | 6 | ||
| d | Stylet short (<22µm) | 7 | ||
| 6 | a | Cuticle with longitudinal ridges | Campbellenchus | |
| b | Cuticle without longitudinal ridges | Gracilancea | ||
| 7 | a | Head with setae | 8 | |
| b | Head without setae | 9 | ||
| 8 | a | Vulva covered by longitudinal flap; male without caudal alae; male cloaca with hypoptygma | Atylenchus | |
| b | Vulva with lateral flaps; male with caudal alae; male cloaca raised | Eutylenchus | ||
| 9 | a | Cuticle with longitudinal ridges | 10 | |
| b | Cuticle without longitudinal ridges | 14 | ||
| 10 | a | Cone about 1/3 of stylet length | 11 | |
| b | Cone almost 1/2 of stylet length | 13 | ||
| 11 | a | Transverse striations not visible through longitudinal ridges | Basirienchus | |
| b | Transverse striations and longitudinal ridges form block (tessellate or areolate) pattern; lateral field with 4 lines | 12 | ||
| 12 | a | Lip region with 2-3 annules, stylet without knobs | Neothada | |
| b | Lip region with 6-7 annules, stylet without knobs | Basirienchus | ||
| 13 | a | Vulva covered by longitudinal flap; stylet 17-19µm | Pleurotylenchus | |
| b | Vulva with lateral flaps; stylet <15µm | Coslenchus | ||
| 14 | a | Cone about 1/3 of stylet length | 15 | |
| b | Cone almost 1/2 of stylet length | 28 | ||
| 15 | a | Head high, with distinct lateral amphid slits | 16 | |
| b | Head variously shaped, amphid slit longitudinal | 19 | ||
| 16 | a | Female body ventrally curved or spiral; female with offset spermatheca and oocytes in multiple rows | Boleodorus | |
| b | Female body straight; oocytes not in multiple rows | 17 | ||
| 17 | a | Tail bent or hook-shaped near tip | Basirienchus | |
| b | Tail more or less straight | |||
| 18 | a | Stylet without knobs, anterior part with wide lumen | Neopsilenchus | |
| b | Stylet with or without knobs, anterior conical with narrow lumen | Basiria | ||
| 19 | a | Head with disc-like structure | 20 | |
| b | Head with smooth contour | 22 | ||
| 20 | a | Head with small disc | 21 | |
| b | Head with large dome-shaped structure | Cucullitylenchus | ||
| 21 | a | Very slender (a=62-76); caudal alae concave posteriorly | Mitranema | |
| b | Less slender; caudal alae rounded | Filenchus | ||
| 22 | a | Very slender (a=60-180); caudal alae lobed | 23 | |
| b | Body width variable; caudal alae rounded if present | 24 | ||
| 23 | a | Head quadrangular; pore-like amphid apertures; body constricted after vulva | Ecphyadophora | |
| b | Head flattened; long amphid apertures; body not constricted after vulva | Ecphyadophoroides | ||
| 24 | a | Cuticle deeply incised | 25 | |
| b | Cuticle not deeply incised | 26 | ||
| 25 | a | Head quadrangular; body striations with zigzag pattern; male without caudal alae | Miculenchus | |
| b | Head flattened; male with caudal alae | Malenchus | ||
| 26 | a | Very slender; indistinct striation; head very flat; long, sinuous amphid aperture | Lelenchus | |
| b | Larger body diameter; distinct striation; head quadrangular; aperture not sinuous | 27 | ||
| 27 | a | Head high with longitudinal amphid apertures lateral; clavate stylet knobs; DEGO > 1/2 stylet length behind knobs | Irantylenchus | |
| b | Head quadrangular; distinct striation; round stylet knobs; DEGO < 1/2 stylet length behind knobs | Filenchus | ||
| 28 | a | Vulva with lateral flaps | 29 | |
| b | Vulva without flaps | 30 | ||
| 29 | a | Lateral field of 2 lines; vagina thin; post-vulval sac short | Allotylenchus | |
| b | Lateral field of 3 lines; vagina thickened; post-vulval sac short | Aglenchus | ||
| c | Lateral field of 4 or 6 lines; vagina not thickened; post-vulval sac long | Cephalenchus | ||
| 30 | a | Lateral field and striations inconspicuous; caudal alae very small | Polenchus | |
| b | Lateral field and striations distinct; caudal alae distinct | Tylenchus |
Commonly occurring in most soils. Feed on algae, mosses, lichens and plant roots. As an example: Soil nematodes were studied in three spruce forests in the Czech Republic from 1988 to 1991. A total of 74 species occurred, most belonged to the orders Tylenchida, Rhabditida and Dorylaimida. The most abundant nematodes were the mycophytophagous species of the family Tylenchidae followed by bacteriophages, especially by those in the order Rhabditida.
Members of the Tylenchidae frequently constitute up to 30% of the individuals in a soil sample.(Ferris and Bongers, 2006)
Probably fairly small. Most reports are about occurrence and abundance rather than documenting any effects on growth. Arguably, the benefits of soil food web stimulation and participation in nutrient cycles may outweigh any economic damage to crops.
Ectoparasites of plant roots, root hairs, algae, etc.
Yeates et al. (1993a): placed the Tylenchidae in the following feeding groups:
Aglenchus: plant feeder (epidermal cell and root feeder),
Cephalenchus: plant feeder (ectoparasite),
Coslenchus: (epidermal cell and root feeder),
Filenchus: (epidermal cell and root feeder),
Malenchus: (epidermal cell and root feeder),
Tylenchus: plant feeder (algal, lichen (algal or fungal component), or moss feeders that feed by piercing), or hyphal feeder (?).
Yeates et al. (1993b): classified Tylenchus spp. as "plant associated", indicating that they were found in the rhizospheres of plants.
Okada et al (2002 and 2003) have demonstrated that Filenchus misellus is a fungal-feeding nematode and able to reproduce on a range of fungi.
Nematodes in this family are associated with algae, mosses, lichens, fungi, and plant roots. Some of the species are considered as weak parasites of plants (Siddiqi, 2000).
The following is an initial compilation of feeding habits of Tylenchidae by Erik Schaper while working on an MSc-project under the supervision of Tom Bongers (Laboratory of Nematology, Wageningen University)..
Project Title: "Tylenchidae, plant parasites or fungal feeders?".
Objectives:
Literature Review:
Additionally:
Okada et al (2002 and 2003) have experimentally demonstrated that Filenchus misellus is a fungal-feeding nematode and able to reproduce on a range of fungi.
Little is known of the life history of most species, but in so far as known, they have no specialized cycles, resting, or resistant stages.
Relatively slight, small stylets penetrating only thin cell walls.
Andrássy, I (1976), Aglenchus costatus, C.I.H. Descriptions of plant-parasitic nematodes, set 6, No. 80, 2 pp.
Baujard, P. (1995), Laboratory methods used for the study of the ecology and pathogenicity of Tylenchida, Longidoridae and Trichodoridae from rainy and semi-arid tropics of West Africa, Fundamental and Applied Nematology, 18, 63-66
Cobb, N.A. (1925), Biological relationships of the mathematical series 1, 2, 4, etc., Chapter 15 in: Contributions to a Science of Nematology,
Ferris, H., and Bongers, T. 2006. Nematode indicators of organic enrichment. Journal of Nematology 38:3–12.
Ferris, H., Venette, R.C., Lau, S.S. (1996), Dynamics of nematode communities in tomatoes grown in convential and organic farming systems, and their impact on soil fertility, Applied Soil Ecology, 3, 161-175
Geraert, E. 2008. The Tylenchidae of the world: identification of the family Tylenchidae (Nematoda). Gent, Academia Press: 540 pp
Geraert & Raski (1987) Rev. Nematol. 10(2):143-161.
Gowen, S.R. (1970), Observations on the fecundity and longevity of Tylenchus emarginatus on sitka spruce seedlings at different temperatures, Nematologica, 13, 267-272
Hanel, Ladislav. 1996. Comparison of soil nematode communities in three spruce forests Boubin Mount, Czech Republic. Biologia (Bratislava) 51.
Hooper, D.J. (1974), Cephalenchus emarginatus, C.I.H. Descriptions of plant-parasitic nematodes, Set 3, No. 35, 2 pp.
Khera, S., Zuckermann, B.M. (1962), Studies on the culturing of certain ectoparasitic nematodes on plant callus tissue, Nematologica, 8, 272-274
Khera, S., Zuckermann, B.M. (1963), In vitro studies of host-parasite relationships of some plant-parasitic nematodes, Nematologica, 9, 1-6
Micoletzky, H. (1925), Die freilebenden Süsswasser- und Moornematoden Dänemarks nebst Anhang über Amöbesporidien und andere Parasiten bei freilebenen Nematoden, D. Kgl. Danske Vidensk. Selsk. Skrifter, Naurvidensk. og Mathem., ser 8, 10, 57-310
Okada, H., Tsukiboshi, T., Kadota , I., 2002. Mycetophagy in Filenchus misellus (Andrássy, 1958) Raski & Geraert, 1987 (Nematoda: Tylenchidae), with notes on its morphology. Nematology 4, 795-801.
Okada, H., Kadota, I., 2003. Host status of 10 fungal isolates for two nematode species, Filenchus misellus and Aphelenchus avenae. Soil Biology and Biochemistry 35, 1601-1607.
Qing, X., Bert, W. 2017. Redefinition of Genus Malenchus Andrassy, 1968 (Tylenchomorpha: Tylenchidae) with Additional Data on Ecology. J. Nematology 49:189-206.
Siddiqi, M.R. (1986), Tyenchida, parasites of plants and insects, CAB, Slough, 645 pp.
Siddiqi, M.R. 2000. Tylenchida, Parasites of Plants and Insects, 2nd ed. Wallingford, UK: CAB International, 833 pp.
Sutherland, J.R. (1967), Parasitism of Tylenchus emarginatus on conifer seedling roots and some observations on the biology of the nematode, Nematologica, 13, 191-196
Thorne, G. (1961), Tylenchinae, chapter 5 in: Principles of Nematology, McGraw-Hill Book Company Inc., New York - Toronto - London, 553 pp.
Wood, F.H. (1971), Studies on the biology of soil-dwelling nematodes from tussock grassland, Ph.D. thesis, University of Canterbury, New Zealand, 286 pp.
Wood, F.H. (1973a), Life cycle and host-parasite relationships of Aglenchus costatus (de Man, 1921) Meyl, 1961 (Nematoda, Tylenchidae), New Zealand Journal of Agricultural Research, 16, 373-380
Wood, F.H. (1973b), Nematode feeding relationships, feeding relationships of soil-dwelling nematodes, Soil Biology and Biochemistry, 5, 593-601
Yeates, G.W., Bongers, T., Goede, R.G.M. de, Freckman, D.W., Georgieva, S.S. (1993a), Feeding habits in soil nematode families and genera - an outline for soil ecologists, Journal of Nematology, 25 (3): 315-331
Yeates, G.W., Wardle, D.A., Watson, R.N. (1993b), Relationships between nematodes, soil microbial biomass and weed-management strategies in maize and asparagus cropping systems, Soil Biology and Biochemistry, 25, 869-876